![]()
Biological Clocks in Mosquitoes
|
 A largely littoral European species, type locality
Bolsward, Friesland, Holland, which is found between latitudes 35°N and
55°N (Marshall, 1938; Stone et al., 1959). According to White (1978) it
is a Palaearctic member of the Anopheles maculipennis complex.
A largely littoral European species, type locality
Bolsward, Friesland, Holland, which is found between latitudes 35°N and
55°N (Marshall, 1938; Stone et al., 1959). According to White (1978) it
is a Palaearctic member of the Anopheles maculipennis complex.
Experimental material
The adults were reared, under LD 12:12, from eggs obtained from the
Ross Institute of Tropical Medicine, where a colony originating from
Holland (latitude around 52°N) had been maintained since 1962. This
colony was a mixture of two strains showing differences in behaviour in
the presence of DDT insecticide (Gerold & Laarman, 1964).
Experimental regimes
LD 8:16 to LD 4:20, nine females, studied from 3 days
post-emergence (4 October 1968). Recorded in LD 8:16 for days two to
five then light-on delayed 4h to give LD 4:20 until day eight.
LD 12:12 to LD 15:9, six females, studied from 3 days
post-emergence (4 October 1968). Recorded in LD 12:12 for days two to
four then light-on advanced 3h to give LD 15:9 until day six.
LD 20:4, eight females, studied from 5-6 days post-emergence (10
October 1968) and recorded for days four to six.
Results and discussion
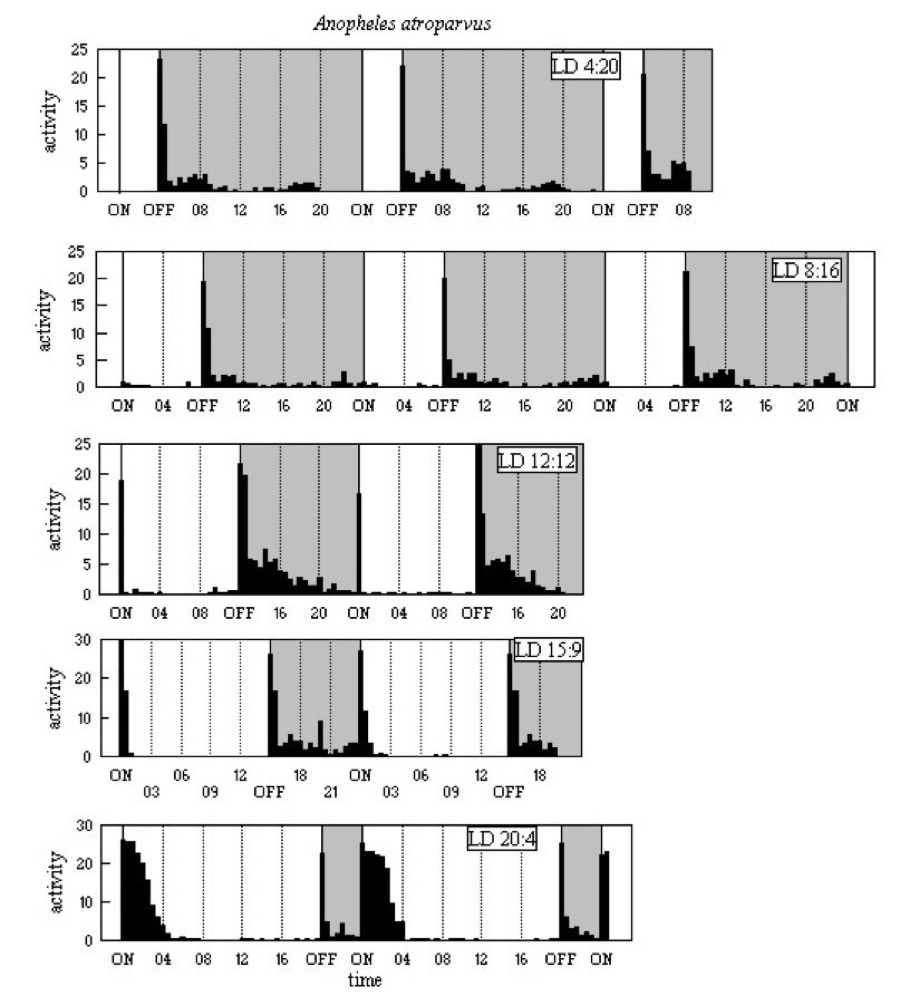
The activity patterns are shown in Figure A6 below. In all five LD
regimes there is a major E peak, with a decline in activity
over the next 3.5-4h. M peaks are clear at light-on in LD 12:12
and LD 15:9, and there is low level activity around 13-14h after
light-off in LD 8:16 and LD 4:20. The major peak of activity from
light-on in LD 20:4, with a high level for the next 2h and continuing
but declining activity for a further 2h, also suggests something more
than just a startle response to light-on. The laboratory pattern of
activity in the summer daylengths, L 16-18h, of its reported
distribution fits well with the pattern most often observed for Anopheles
malaria vectors; that is, post-dusk to near midnight feeding activity
and dawn egress from human habitations.
There does not, however, seem to be any specific field report of activity timing for An. atroparvus. Lachmajer (1952) reported that, near Danzig, Poland (54°22'N), overwintering females became active in the open in late May and the July population consisted of old females that had oviposited several times. Adults of the first, summer, generation emerged in increasing numbers from the end of August onwards, with emergence continuing until November. The females of the second generation do not oviposit until the next Spring and very few summer females survive the Winter. The bulk of the hibernating population is of the autumn generation. Similar findings have been found in southern England, where Ramsdale & Wilkes (1985) collected overwintering females in animal shelters. Re-activation took place in March-April, leading to a new generation of adults in mid-June. They noted evidence for intra-specific geographical variation and considered that, given the considerable differences in duration of Winter within its distribution, regional differences in reaction to daylength could be expected.
Figure A6
![]()
|
©1998, 2010 - Brian Taylor CBiol FSB FRES 11, Grazingfield, Wilford, Nottingham, NG11 7FN, U.K. Comments to dr.b.taylor@ntlworld.com |
href="\crhtml\anatro.htm"