Biological Clocks in Mosquitoes - Section 2
|
(i) Phase response evidence and phase reference points
The definitions of "phase response curves" and "phase reference points" have been mentioned briefly earlier, drawing on Friesen et al. (1993), but the whole question is worth exploring.
Page (1991) examined the circadian pacemaker of the cockroach and the relation between pacemaker period and response to light. He used onset of activity as the phase reference point and light onset as the reference for the pulse, but why these points were chosen was not clear (perhaps it was simply an arbitrary choice).
Moore & Rankin (1993) examined the walking rhythm of honeybees, Apis mellifera ligustica, to try and determine if entrainment could be accomplished by the actions of discrete signals, such as light-on and light-off, or single pulses of light at the normal light-dark transition points. Their exploration commenced with the reasoning of DeCoursey (1972) that little or no phase change should occur for a wide variety of LD ratios. The bees were exposed to a range of LD regimes, from LD 4:20 to LD 22:2, and the expectation was that under conditions of discrete entrainment the phase of the rhythm would maintain a fixed position to with respect to light-on or light-off or a position indicating a proportional effect between the two signals. They tested their results for correlation between three "phase reference points" (activity onset, peak activity and activity offset) and light-off. When the pattern in LD 22:2 was included only the activity offset showed a significant correlation with light-off. When the LD 22:2 results were omitted from the statistical test, peak activity and, to a lesser extent, activity onset also were shown to be correlated with light-off. Light pulse experiments, using a single hour of light either following the original light-on or preceding the original light-off in a preceding LD 8:16 regime, had shown that the rhythm remained fixed when light-off remained unchanged but advanced when light-off was seven hours early. Two limitations, however, can be placed on Moore & Rankin's findings.
Figure 53 shows a photoperiodogram extrapolated from their
results
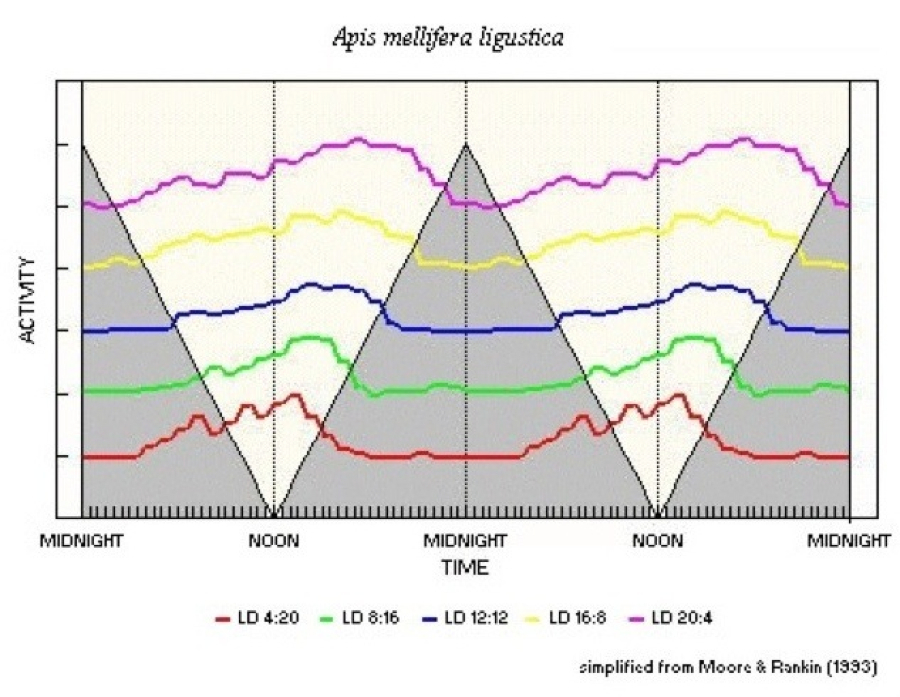
First, activity offset could be due more to inhibition of activity by the lack of light, as their short LD results showed that activity will resume some 14h after light-off. Second, the activity was unimodal with little variation in activity level from noon to light-off in LD 12:12. The LD 22:2 result could be interpreted as showing that activity builds up from light-on to a hump peaking around 13-14h later and then declines smoothly, with the "real" minimum being some 18-20h after light-on.
If the argument for multiple clocks is accepted then the whole issue of phase response curves is cast into doubt. Perhaps most importantly, the clocks are not fixed in relation to each other. If they were then no possibility would exist for adjustment to meet the changing day-lengths experienced as the seasons progress. The elegant argument made by Dreisig (1980) for an endogenous cycle of sensitisation can be questioned on this ground, as he made no reference to changing daylengths. The mosquito evidence shows also that all light pulses, because they cross both the upward and downward light thresholds, may have a double entraining effect. Two light pulses, as used in the classical Pittendrigh studies, would mean a double-double entrainment. If a single short duration pulse is considered, what seems likely is that the threshold transition will shift, or entrain, a particular sine wave by no more than a certain proportion depending on when the pulse falls. Examination of the sine wave form (for example see Figure 28) shows how the rate of change, or slope, of the wave is far less around the apogee and perigee than in mid-cycle. Thus, taking the assumption of a light-threshold transition as producing a 5% shift in the direction of the closest apogee or perigee and a simple t = 24h, such a shift could return the wave to the maximum or minimum for some two hours either side of the apogee or perigee; but in mid-cycle a 5% shift would encompass less than 30 mins and thus engender a minimal "phase response".
Smith (1983), for Lucilia cuprina, used a 15 min light pulse in DD following LD 12:12, and his phase response curves reveal precisely such a pattern of adjustment. L. cuprina is a fully-day-active species and thus may differ from almost all the mosquitoes which have been studied but the phase-response curve defined by Jones et al. (1972) for the late crepuscular mosquito An. gambiae also shows the phase shift to be least in mid-cycle (around 6h or 18h after the original light-off). To return to the L. cuprina results, Smith (1983) gave histograms of the effect of extending the last light period from 12h to 20h and 30h, before entering DD. He used a simple single sine wave to extrapolate how the phase is reset and argued that the peak (of an assumed unimodal rhythm) shifts by no more than 12h and then always follows the last light-off by 15h (two-thirds of t = 22.5h). Close examination shows that the activity actually formed peaks with a different shape, depending on the duration of the final L. If one argues that the apparently unimodal activity, normally effectively filling all daylight hours in LD 12:12, is the manifestation of at least two clocks (on- and off-entrained) then extension of the last L will alter the relative conjunction of the clock effects. Moreover, the L. cuprina activity histograms showed deviations from a smooth sine form which are consistent with the multi-clock concept. Finally, discussion of rhythms of single once-only events such as pupation and eclosion, is limited as the underlying mechanisms cannot be observed on a repetitive basis for individuals, and, indeed, all the findings are based on populations.
(ii) Photoperiodism
The sensitivity of most insects to low temperatures has led most species of temperate latitudes to adopt an overwintering survival technique. The phenomenon known as diapause, in which short-lived species have more than one generation during the Summer and then switch to production of a generation which remains in a state of arrested development (usually in the egg or early instar stage), has been the subject of extensive research. On balance, current thinking (see Dumortier, 1994; and Vaz Nunes, 1994, for recent reviews) seems to be that the "photoperiodic clock" does involve a circadian system. A key question remains as to how a circadian system can provide a means of time measurement. It may be unwise to proffer the evidence from studies of mosquito flight activity as a contribution to the photoperiodism debate but justification can be found in Dumortier's paper. There a methodological difficulty in testing the involvement of the circadian system was expressed as there being no concrete parameters related to the photoperiodic clock running whose change in time can be measured (Dumortier 1994). The new concept, with its separation of clocks and variable Lt, offers a simple method of assessing the duration of daylength by coincidence or otherwise of the members of each pair of clocks.
Alternatively, if the effects of the clocks are summative (see Figure 52) the degree of coincidence could mean sufficient diapause inhibiting factor or vice-versa. The argument for time measurement based upon imbalance between the clocks, moreover, bears a resemblance to the system of photoperiodic time measurement known as "internal coincidence", the simplest version of which was described by Dumortier (1994) as first being expressed by Pittendrigh (1972). This argued for two oscillations, A and B, phase-set by light-on and light-off respectively, thus assuming a phase relationship which changes with varying night- and day-lengths. This suggestion was predated by the demonstration of light-on and light-off entrainment in the papers on Ae. aegypti flight activity (Taylor & Jones 1969) and the outline paper relating it to geographical range (Taylor 1969b).
Interestingly, Dumortier (1994) cast doubt on the classic experimental techniques known as the "Nanda-Hamner" and "Bunsow" protocols, which use unnatural dark periods, with light pulses in the latter protocol, to investigate the induction of diapause. As the attempts to interpret results from use of such protocols have all assumed only a single clock, a valid multi-clock system must add to the doubts. A final point upon which a valid multi-clock system would cast doubt is the proposal that the internal coincidence, or coupled oscillator, system is composed of a master "pacemaker" and a "slave" (Vaz Nunes,1994). The mosquito evidence indicates that each of the clocks may play an active and determinate role, and the role not only can be separated from those of the other clocks but will remain separate in constant conditions (see, for instance, the activity of Ae. aegypti in LD 20:4, Figure 30, and in LL following LD 20:4, Figure 31a).
END
|
©1998, 2010 - Brian Taylor CBiol FSB FRES 11, Grazingfield, Wilford, Nottingham, NG11 7FN, U.K. Comments to dr.b.taylor@ntlworld.com |