Biological Clocks in Mosquitoes
|
The species was formally designated as Culex pipiens quinquefasciatus Say by Sirivanarkan & White (1978). Numerous other names have been used but the commonest synonym is Culex pipiens fatigans Wiedemann, which can be found in most Old World publications before 1978. Stone et al. (1959) described it as cosmotropical, and its type locality was the East Indies (presumably Indonesia). In the USA, it is found up to 40-42°N, but adults do not appear in large numbers until July or August at these northern limits, and they disappear in September or early October, except for hibernating adults. In the southern USA adults and larvae can be found throughout the year.
Experimental material
The adults were reared, under LD 12:12 or LD 4:20, from eggs obtained
from the Ross Institute of Tropical Medicine, where a colony
originating from Lagos, south-western Nigeria (latitude 6°27'N) was
maintained. For simplicity the present results are designated as LAGOS,
and the Sussex researchers' results (see below) as GAMBIA.
Experimental regimes
LD 4:20, six females, reared in LD 4:20 and studied from 4-5
days post-emergence (5-6 March 1969). Recorded for days two to four.
LD 8:16, five females, reared in LD 4:20 and studied from 4-5
days post-emergence (5-6 March 1969). Recorded for days two to four.
LD 12:12, nine females, reared in LD 12:12 and studied from 4-5
days post-emergence (5-6 March 1969). Recorded for days three to five.
LD 16:8 to LD 20:4, six females, studied from 4-5 days
post-emergence (5-6 March 1969). Recorded in LD 16:8 from days three to
five then light-on advanced by 4h to give LD 20:4 until day nine.
Results and discussion
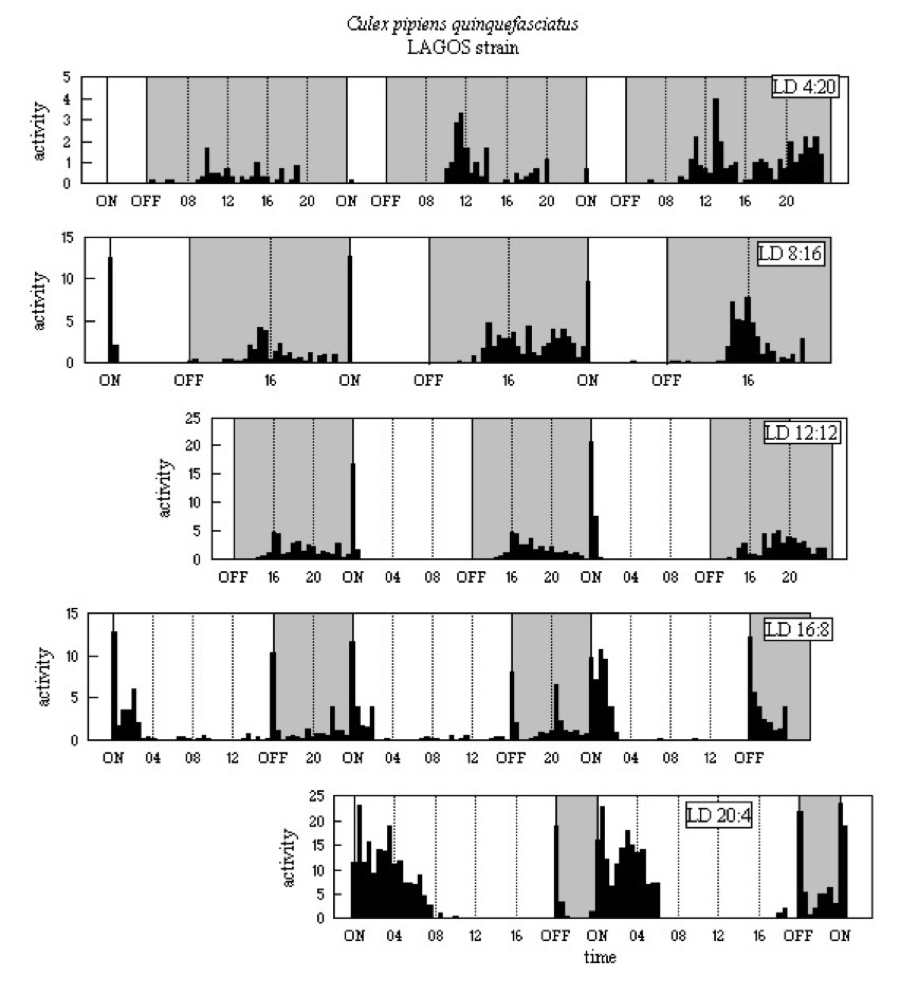
The activity patterns are shown in Figure A12. In all five LD regimes
there is a major N peak, with greatest activity some 6h after
light-off. In the mid-length regimes, LD 8:16 to LD 16:8, a sharp M
reaches the highest level of the whole 24h-cycle; but there is a
broad M running well into the light in LD 20:4. There is E
activity in LD 16:8 and LD 20:4. The activity of LAGOS in LD 12:12
clearly is bimodal with N, some 4-6h after light-off, and a much more
pronounced M, immediately following light-on.
Figure A12
The LAGOS results compare well with Cx. p. quinquefasciatus
activity cycles in field conditions. Service (1963) found at Kaduna,
Northern Nigeria (10°30'N), that biting occurred throughout the night,
peaking between 2300h and 0100h. Teesdale (1960) used window traps at
Mombasa, Kenya (4°4'S), and found peak exit flight at 1800-2100h and
0700-0800h, biting took place between 2100h and 0600h but peaked
between 0100-0300h. Mating in the laboratory was observed by Sebastian
& De Meillon (1967) to start early in the evening, with a first
peak at around 2000h and a slight fall before a maximum at 0400-0600h.
Similar activity results were found also by Kaul & Wattan (1978),
who studied swarming and mating behaviour in the field at Delhi, India.
The LAGOS pattern, however, differs from the pattern for GAMBIA, in which the major activity of the 24h-cycle was an E peak, occurring immediately after light-off (see Jones, 1976, for the first report). Although attention was drawn to the absence of E in LAGOS (Taylor, 1977), this difference appears to have gone unregarded by the Sussex researchers. This is despite the fact that the sophisticated interpretation of the circadian mechanism exhibited by GAMBIA has been based solely on E being a "reliable phase marker" (Peterson, 1980a, b, c). What has emerged from investigations of virgin and inseminated GAMBIA females (Jones & Gubbins, 1979) is that N became manifest after insemination. M in virgins was shown as a build-up from 5-6h before light-on, with a sharp response to light-on; but M in inseminated was solely a sharp burst of activity following light-on. E differed little. The early GAMBIA results were acknowledged to be with virgin females and males. As the LAGOS females had emerged and been held in a large cage together with males from the same egg batch, it is likely that the LAGOS results are for inseminated females. As noted above, the N peak in both LAGOS and GAMBIA inseminated females matches the many field reports of peak biting activity.
The wider significance of the patterns of activity in the other LD regimes will be discussed elsewhere but comparison can be made with the results of Jones (1982), who showed patterns of activity for both virgin and inseminated GAMBIA females, recorded in LD 6:18 and LD 18:6. Although LAGOS shows an E peak in both LD 16:8 and LD 20:4, this could be due to a simple forward shift of the N peak, as a similar shift occurred in GAMBIA. The pattern shown by LAGOS in LD 4:20 and LD 16:8, which is a build-up of activity towards an M peak preceding light-on (perhaps more truly an M' peak), actually resembles the GAMBIA pattern for virgins rather than inseminated females. Similarly, in LD 16:8 and LD 20:4, the amplitude and breadth of the LAGOS M peak more closely resemble the activity of GAMBIA virgins in LD 18:6.
|
©1998, 2010 - Brian Taylor CBiol FSB FRES 11, Grazingfield, Wilford, Nottingham, NG11 7FN, U.K. Comments to dr.b.taylor@ntlworld.com |