Biological Clocks in Mosquitoes - Section 2
|
Figure 11
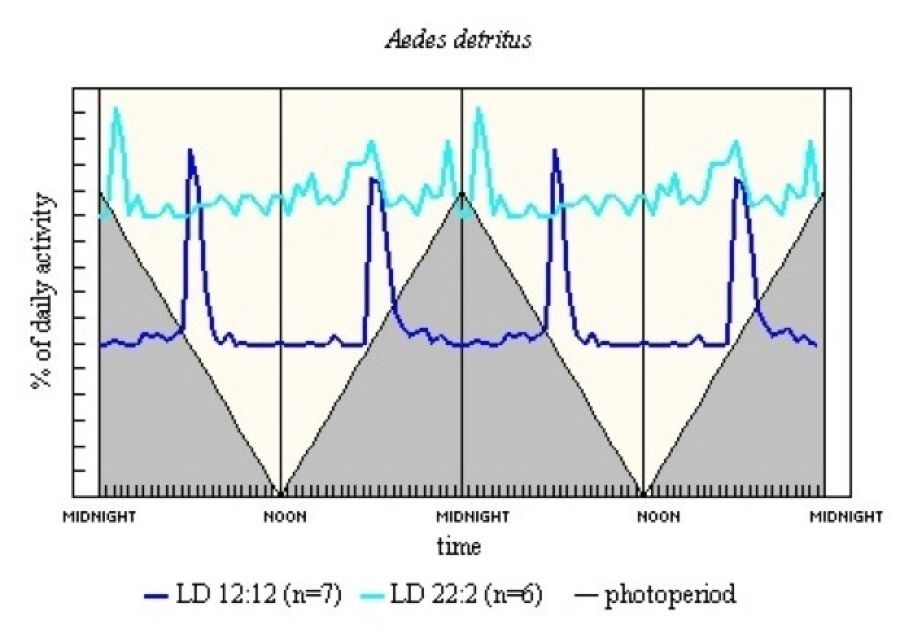
Figure 12
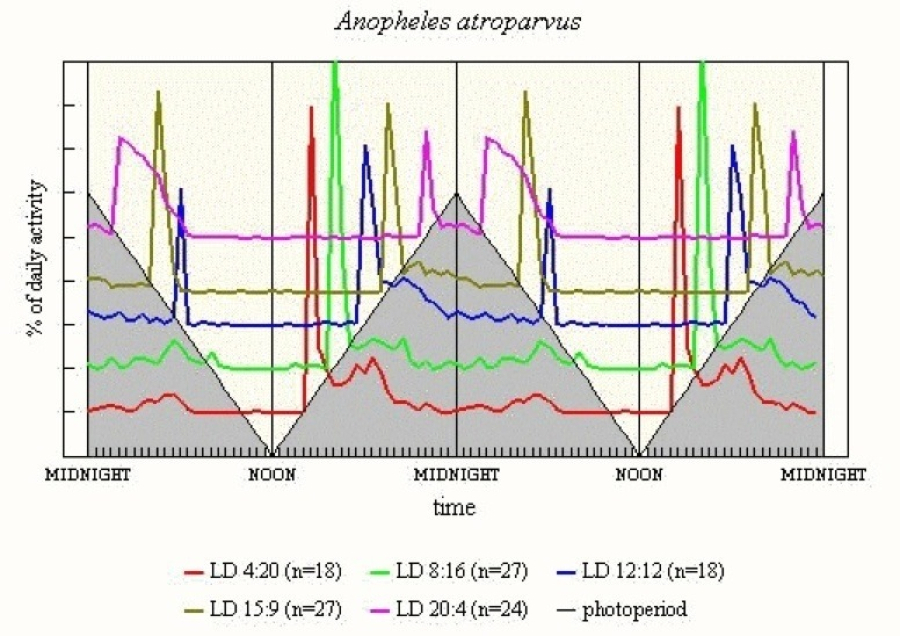
Figure 13
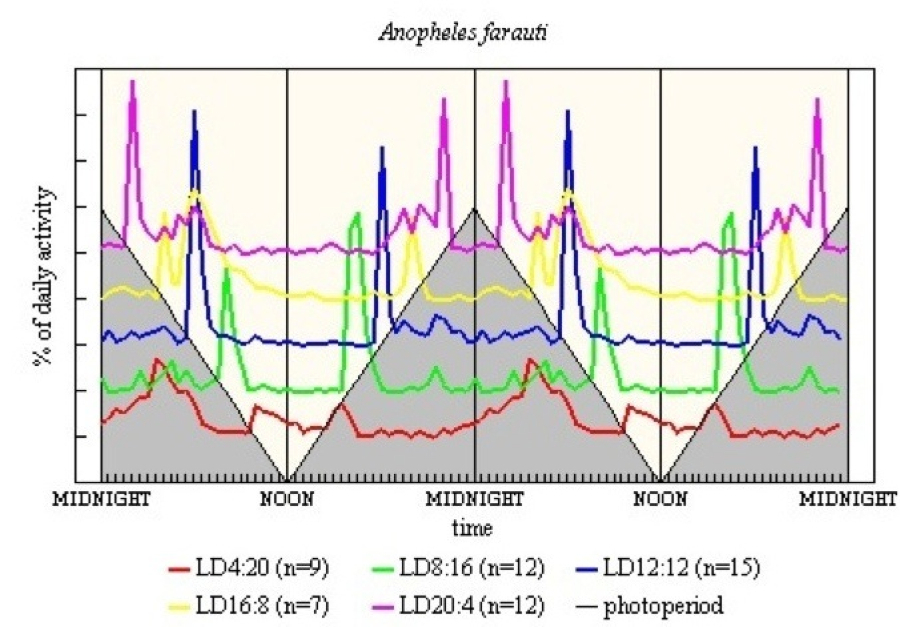
Figure 14
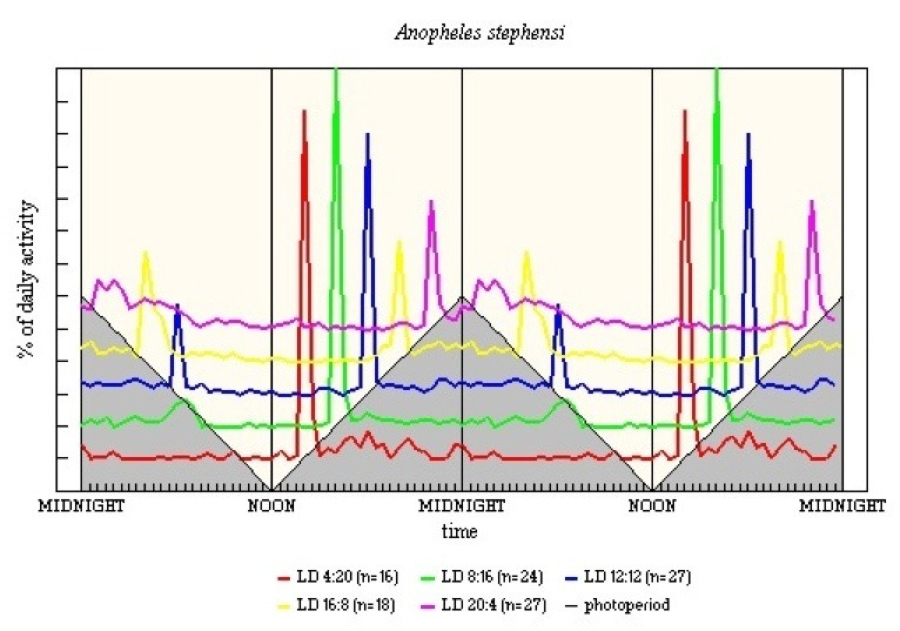
Figure 15
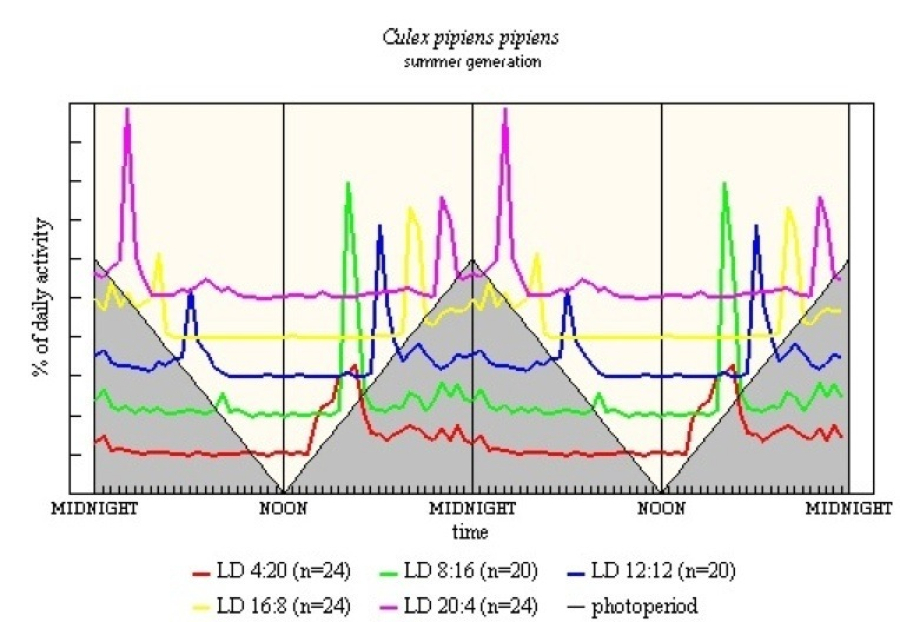
Summer Generation - reared from larvae collected when the natural LD was 16:8. Figure 16 shows the photoperiodogram. In all five LD regimes there is a major E peak, with moderate activity throughout the night, except when L < 12h when activity ceases some 8-10h after light-off. The E peak is more diffuse and somewhat late in LD 4:20, and there is some activity before light-off in LD 20:4. M peaks are clear at light-on in the three long L regimes. Diffuse peaks occur some 8-10h after light-off in LD 4:20, LD 8:16 and LD 20:4. The last is the only regime in which significant activity occurs in the light.
Figure 16
Hibernating Generation - adults collected in January, natural daylength about 8h 30m. Figure 17 shows the photoperiodogram. In LD 8:16 the M activity is greater than E. When L = 12h or longer, similar levels of E and M activity formed a clear bimodal pattern. Entrainment of E by light-off some 24-27h before would explain the apparent single peak encompassing the dark period in LD 20:4; and also why there is a midnight burst of activity in LD 4:20. The most obvious differences between the summer and winter, hibernating, generations is that in the short L regimes the latter are less active but have a more marked M.
Figure 17
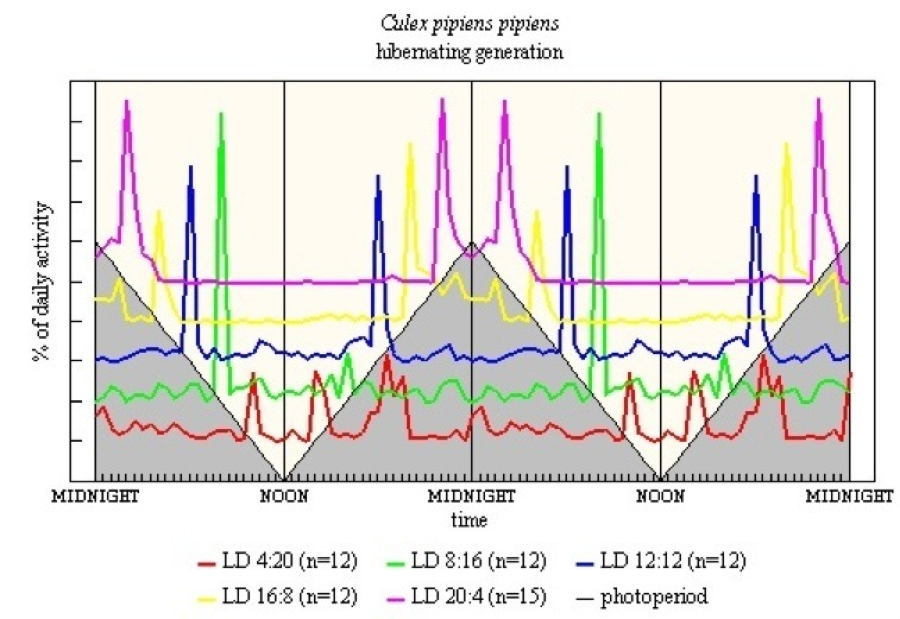
Figure 18
The Ae. detritus results (see Figure 11) show a very clear E and M pattern in LD 12:12. In LD 22:2 there is a peak some 16.5h after light-on, which suggests light-on may have an entrainment effect.
An. atroparvus (see Figure 12) shows pronounced E, with activity continuing for some 5-6h when L < 16h. M activity is sharp in LD 12:12 and LD 16:8, but a small peak can be seen some 14-15h after light-off in the short L regimes. The broad M in LD 20:4 perhaps can best be explained as a resurgence of the E activity. Light clearly has a strong inhibitory effect on activity.
With An farauti (see Figure 13) the main difference is that the entrainment of M seems to be by light-off 11-13h before, this can be seen as split M peaks in all except LD 12:12. E also is imprecise or split in the long L and long D regimes.
The light-off entrainment of M in An. stephensi appears to be within a wider framework of 8-15h (see Figure 14); however, M is rather low level and is not seen in LD 4:20. E activity is more closely confined, being restricted to the first hour after light-off, except in LD 4:20.
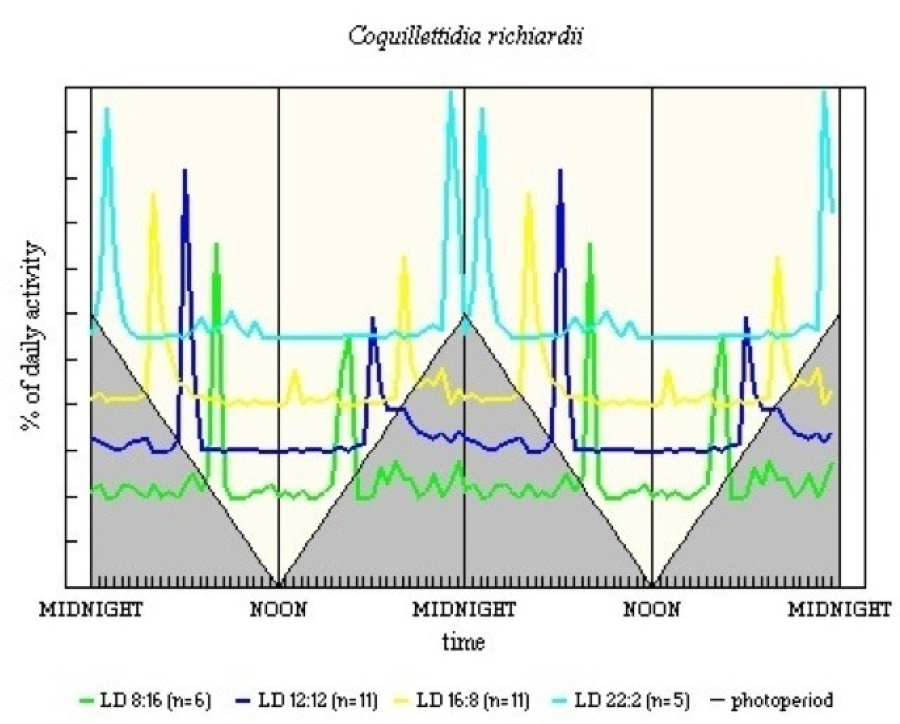
The relatively limited results for C. richiardii generally conform to the pattern (see Figure 15). The major peak around the very short D in LD 22:2 seems most likely to be a manifestation solely of E, with inhibition by D, as there is a small diffuse M 10-11h after light-off, which is well into the light period.
With Cx. p. pipiens both summer (see Figure 16) and hibernating (see Figure 17) generations were studied. The summer generation shows diffuse E in LD 4:20, with the peak being 3h after light-off. In LD 20:4 there is early E and late M. The hibernating generation shows a rather different pattern. This is influenced partly by the low level of activity in the short L regimes (see Appendix); activity in LD 12:12 is much more precise and more pronounced. The LD 20:4 pattern for both generations shows similarity with C. richiardii, again perhaps there is a double manifestation of the E peak.
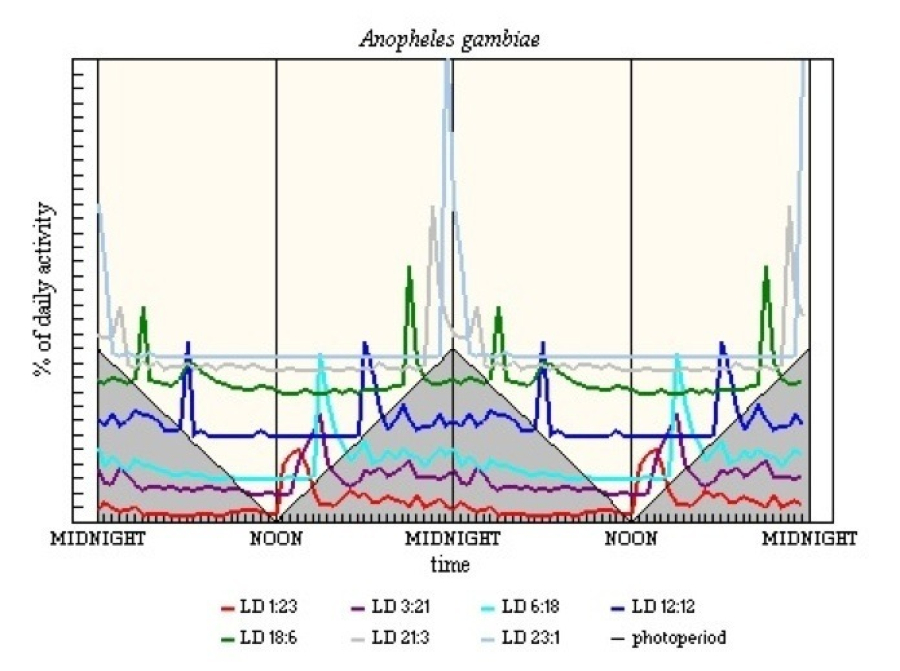
The pattern of activity in An. gambiae is similar to the other late-crepuscular anophelines (see Figure 18). The LD 23:1 and LD 21:3 results fit the suggestion of a bifurcation of E activity and M being some 12h after light-off. The strong M in middle LD regimes was regarded by Jones et al. (1972) as a startle response to abrupt light-on, but the evidence, especially the peak well into L in LD 18:6, does allow for there being a genuinely entrained M.
Additional evidence of a mosquito in this group can be found in Nayar & Sauermann (1971). They exposed Aedes taeniorhynchus to a range of LD regimes and found it to be a nocturnal species, with a clear bimodal E and M pattern in LD 12:12, which persisted in DD after LD 12:12. In regimes with D > 12h the M peak settled around 12-15h after dusk. In L > 12h activity compressed into D, with light inhibiting activity, except in LD 23:1 when there was activity starting some 12h after light-on. They did not comment on the drifting of peaks away from the bimodal E and M. The East Asian species, Culex pipiens pallens, also shows this group pattern in LD 16:8 and LD 12:12 (Chiba, 1964; Chiba et al., 1982).
 NEXT
NEXT
|
©1998, 2010 - Brian Taylor CBiol FSB FRES 11, Grazingfield, Wilford, Nottingham, NG11 7FN, U.K. Comments to dr.b.taylor@ntlworld.com |