Chapter 4 - Economic Importance of Ants - Ants as Vectors of Plant Disease
| The Ants of
Africa Chapter 4 - Economic Importance of Ants - Ants as Vectors of Plant Disease |
|
| Economic Importance - Introduction |
First observations - The role of ants in the dissemination of the cocoa disease caused by species of fungi of the genus Phytophthora was first suggested by Dade (1927). Over 40 years later, A.M. Gorenz (1970) described how, at Akure in Nigeria in 1968,
"A small black ant also has been implicated as an agent in bringing Phytophthora from the soil to pods on which it attends to scale insects. Two trees with no black pod, at the margins of a cocoa plot had pods above the ground with scales on the pod stalk being cared for by the ants. Two sporulating diseased pods were placed at the base of each tree and covered with litter. Two weeks later all seven pods (three on one tree, four on the other tree) had the black-pod disease starting at the petiole side where the ants had built tents made of soil and debris to protect the scales from the rain. Pods at the same height but without the ant on three adjacent trees remained healthy and free of the disease".
A typical ant-mediated infection and sequence infections arising by drips containing spores is shown in the photograph.
E.K. Okaisabor (1971) continued the work in Nigeria and showed how certain ants, listed as Camponotus species, Crematogaster africana, Tetramorium aculeatum (as Macromischoides aculeatum) and Oecophylla longinoda could transmit the disease. This, however, was demonstrated as an effect of the ants moving across diseased pods with the fungus in the sporulating stage and passively transporting spores on their body surfaces. This led him to investigate the effect of a mixed insecticide-fungicide spray on fungal incidence, achieving a better control than fungicide alone (Okaisabor, 1972).
Harry Evans, 1971, 1973a, 1973b. Contemporaneously in Ghana, Evans carried out similar studies but also examined the effect of tent-building ants. He showed that soil-utilising ants could contribute to vertical spread by transporting contaminated soil upwards from the base of the tree. The commonest of such ants included Camponotus acvapimensis and Pheidole megacephala which he described as frequently occurring around the edges of cocoa farms. A contribution to horizontal spread at the canopy level was shown to be made by Crematogaster striatula, which he found transporting material from diseased pods to utilise in tent-building. Although he noted that other ants had similar habits, they were not listed because, unlike Crematogaster striatula, they were of limited occurrence.
All the foregoing work was solely of a qualitative nature but it formed an important part of the background to the extensive efforts of the International Black Pod Research Project, based at CRIN between 1973 and 1980. In terms of the ant-black pod research, two important facts emerged from the specialist plant pathology studies.
First, Mike Griffin, in collaboration with Clive Brasier, found that two species of the fungus were involved, the well-known Phytophthora palmivora (Butler) Butler and the new species Phytophthora megakarya Brasier & Griffin. This had major significance, as P. megakarya was found to be much the commoner in Nigeria and eastwards, and to be much better at surviving the dry season in a soil phase, possibly in the cocoa root hairs (Brasier et al., 1981).
Second, Alan Maddison made a very detailed study of the fate of all pods on a 16-tree plot, using three-dimensional mapping. This revealed that "debris" ant tents, built by the small Crematogaster (Orthocrema) species 1, could be the source of a disease outbreak at the highest level on the trees, at over 5 m above ground. As infections were differentiated into "initiator" and "sequence" infections, it was possible to show debris tents were the initiators for 46 infections and led to 57 sequence infections (together totalling 10.9% of all infected pods). Soil tents, built by Pheidole megacephala, led to only 9 initiators but a further 57 sequence infections (together totalling 7.2% of all infected pods) (Maddison, 1981).
The E5/1 study, 1975 - The first year of the 3-D mapping was 1974 and the results contributed to the planning of the major study in the CRIN cocoa block E5/1 in 1975 (Taylor & Griffin, 1981).
As a basic routine, all 1139 trees were examined daily for fresh black pod infections by the team led by Mick Griffin. These were classified into initiator infections (with the sources defined as soil and litter splash, ant tents, rodent damage, insect damage, flower cushion and canker, and no obvious source) and sequence infections (which followed from a recognised initiator). Twice weekly all ant tents were recorded and the next day my team of entomology staff ascertained the position and nature of the tent (soil, organic debris or a combination), the tended insects, and the ant species. The ant tent recording was done on 56 days in the season and separate periodic verifications were made together with surveys of other ant species.
When sequence infections (some 60% of all black pods) were ignored, ant tents were found to be the source of around 25% of initiators. However, 46% of the trees had no pods and a further separation of the trees with black pod showed that it was on only some one-third of those that ant tents were the initiators. Although only 18.5 % of the trees had ant tents as initiators, those 215 trees bore 55.5% of all the black pods in the entire experimental area. A key contributory factor was that those trees were also the most productive. In other words, the more pods the tree produced the more the likelihood of there being ant tents; due to there being a greater number of flower cushions with young tissue and thus a greater attractiveness to Homoptera (see also the work in Ghana by Campbell).
E5/1 was a complex site which had been planted as a spacing and thinning trial; the most productive trees were in the areas of the greatest spacing, which accordingly tended to be the areas of lowest canopy density. The results showed clearly how ants could make a major contribution to the annual black pod epidemic. To emphasise this, during April 1975 a total of 58 black pods were recorded in plots studied by the project pathologists. Of those 28 had their origin ascribed to soil splash and 23 to ant tents. Thus, ant activity led to almost 40% of the initial black pods of the seasonal epidemic.
The ant species involved in tent-building at E5/1 were Camponotus acvapimenis, Myrmicaria fumata (as striata), Odontomachus troglodytes, Pheidole megacephala, Crematogaster zavattarii (as Cr. ? gabonensis and Paratrechina grisoni (Para. species 3). The distribution of the ants in the Block is examined in the section on the ant mosaic.
Odontomachus troglodytes tent on pod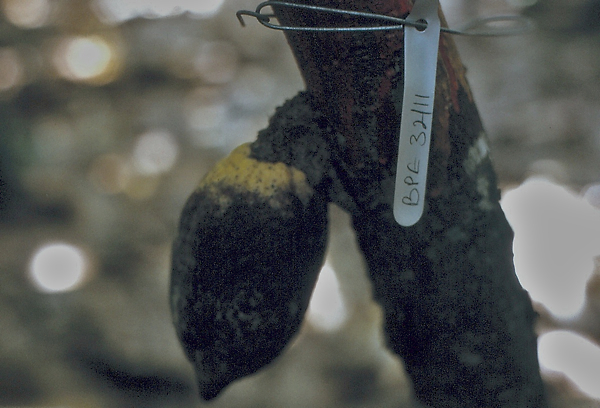 |
Pheidole megacephala tent on pod |
Myrmicaria fumata tent on pod |
|---|
By far the most abundant ant species was Camponotus acvapimensis which was observed at some point in the season building tents on 462 trees. All its tents were built of soil and were found from ground level to 380 cm, on 111 trees the tents were the sources of a total of 264 infections.
Myrmicaria fumata (as M. striata), which was found mostly in association with Camponotus acvapimensis, had soil tents as the sources of 35 black pods on 17 trees.
Odontomachus troglodytes was responsible for 109 infections on only 26 trees. It builds tents of soil removed from its nests among the tree roots and this was the probable cause of the high infection rate, but its capacity to climb is not good and almost all the tents were below 100 cm.
Pheidole megacephala built its tents quite frequently at up to 320 cm, although the higher tents tended to be built of organic debris rather than soil, with the lower soil tents being the main source of 27 black pods on 12 trees.
Crematogaster zavattarii, which is a wholly arboreal species, built carton tents on some 60 trees, most in a distinct single colony, but only two tents were thought to be a source of infection.
Lastly, Paratrechina grisoni was found building tents of organic debris on 14 trees and, despite the findings being on only one or two recording days per tree, four of the tents were sources of black pod.
The number of trees on which ant tents were seen on each of the 56 recording days added up to 6134 tree-days. The insects being found tended by the ants were aphids (Toxoptera aurantii, 91.0%), pseudococcids (2.6%), stictococcids (5.2%) and a combination of two, or three, of these (1.2%). Camponotus acvapimensis was the attendant ant on 64.8% of the tree-days, and also was the attendant for 88.4% of all the stictococcids (most of the remainder being tended by Myrmicaria fumata). Of the pseudococcids, 56% were tended by Camponotus acvapimensis, 20% by Crematogaster nigeriensis and 19% by Pheidole megacephala.
E5/1, 1976-78 - In succeeding years, the E5/1 site was used for testing a range of treatments, such as the insecticide dieldrin and fungicides, to determine their effect on black pod levels (Griffin et al., 1981). The report of the work in 1976, after my own participation had ended, unfortunately, has a mismatch between text and figures so interpretation of the results is not easy. The dieldrin treatment, however, affected not only the ants in the treated plots but also apparently in adjacent plots. This should not have been surprising in view of the colony geography but illustrates the difficulty in demarcating plots of a regular shape on the irregular pattern of ant foraging areas (see the plan of distribution of Camponotus acvapimensis in 1975). The effect of reduced ant activity led to reductions in black pod incidence. Later at CRIN, in 1978 use of insecticide treatments was further studied, on plot N4/1, but without clear results (Ward, Maddison & Adebayo, 1981). The insecticide applied to tree bases certainly stopped the activities of the soil-tent ants but the comparison with the untreated plots showed no reduction in disease. This turned out to be due, at least in part, to the lack of ants in the untreated plots. The studies were extended also to Lafiaji, near Owena, in the wetter Ondo area of eastern Nigeria (1500 mm rain per year, whereas CRIN has about 1000 mm) (Maddison & Idowu, 1981). There, use of tree base insecticide enhanced the effect of fungicide alone in reducing disease incidence.
Also at CRIN in 1978 but apparently without reference to the existing knowledge of ants, Adedeji (1984) attempted to analyse the "nesting boundaries of some dominant ants". As with the 1975 study at E5/1, he made regular (in this case, weekly) records of the "minimum and maximum heights of ant nests along with the number of ant nests, site of ant nests and the ant species involved". It is clear, however, that in using the term "nests" he actually meant "tents". A further error seems to be his claim to have found Crematogaster africana among the ants recorded building the "nests". This the species does rarely, if ever, and I suspect that he was dealing with Crematogaster nigeriensis or some similar Crematogaster (Sphaerocrema) species. He also has Camponotus acvapimensis spelt as Camponotus acyapimensis, and has Camponotus acvapimensis and Myrmicaria striata "nesting" together in the canopy. When the probable corrections are made, his report can be seen to confirm the 1975 findings (Taylor & Griffin, 1981).
Further qualitative work by Babacauh (1982) in the Ivory Coast added strong evidence for the potential of Pheidole megacephala as a vector. He started with a young plantation in which black pod neither had been observed nor detected by mycological techniques. For the first experiment, trees were selected which had ants both nesting at the foot of the tree and actively constructing tents, mainly around the pod stalk and most commonly over the mealybug, Planococcoides njalensis. The main ant was Pheidole megacephala which was building tents of soil and plant debris. Also found building tents were species of Crematogaster and Camponotus. His statement that the Crematogaster ants were building very solid and rain resistant tents using large soil particles, if correct, was a unique finding for any Crematogaster (it seems more likely from the description of the tents that the ant was Myrmicaria striata).
For the experiment, simultaneously the tents were destroyed and a zoospore suspension of a distinctive strain (DIN4) of Phytophthora palmivora was incorporated into the ant nests, together with a red "pate". The tents were reconstructed within 24 h and particles of the pate were seen in the fresh tents, giving a strong indication that the fungal inoculum also had been transported. Ten days later black pod infections were observed around the new tents. Other disease outbreaks occurred on flower cushions and leaf bases, where minute tents had been built over coccids. Finally, 18 black pods, all associated with ant tents and all of the DIN4 strain occurred on eight trees situated within 6 m of the initial placement. Larval Planococcoides njalensis were examined and the ventral side of their bodies were found to be covered in sporocysts of Phytophthora palmivora.
In a further experiment, Babacauh studied six trees, three with nests of Pheidole megacephala at their base and three without tents. Pods growing at mid-tree height were inoculated with DIN4 and the trees were observed for the next four months (July-October 1978). On all trees black pods occurred below the infection sources (17 on the ant-free trees and 8 on the ant trees) but only on the trees with ants were black pods (46 in total) found above the sources. All the infections were of DIN4 and clearly the ants were a significant factor in the upward spread of the disease.
| Economic Importance - Introduction |
©1998, 2003, 2004, 2006, 2012 - Brian Taylor CBiol FSB
FRES 11, Grazingfield, Wilford, Nottingham, NG11 7FN, U.K. |
href="econom2.htm"