![]()
![]()
Deepwater Rice - investigations into the yellow stem borer in Bangladesh - 6 - 1981 research continued |
A side benefit of the regular dissections of stem samples during the two trials was the opportunity to analyze the population structure of the yellow rice borer during each year. Figure 1 shows the composition of the stem borer juvenile population on each date of our observations. Additional points on the graph were derived from two sets of dissections of stem samples from agronomy trials, and from survey data obtained in previous years (where the latter showed a high enough infestation for analysis).
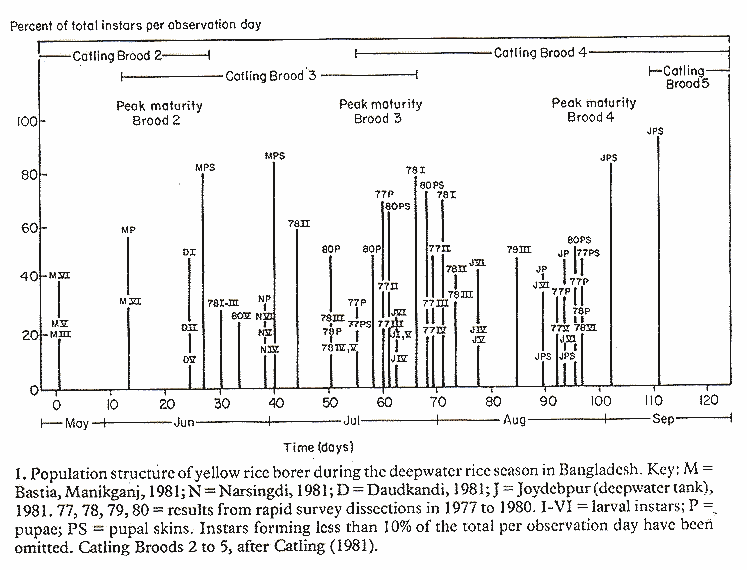
The composition of the population on any one date was expressed as the percentage of the total juvenile stages, which fell into one of the six larval instars, pupae or pupal skins (the last possibly can be deceptive because pupal skins, unlike the other stages, tend to persist as relics). Our first 1981 observations were on 22 May, whereas for 1977-1980 no data were obtained, at least not from the rapid surveys, prior to late June.
In general, our summary of the population structure coincided with the thinking of Catling (1981). The timing of the peak maturity in each brood, we felt might be somewhat earlier than he had suggested, especially of brood 4. The population structure of brood 3 seemed less clear than for broods 2 and 4. Possibly, this was because stem dissections in the middle of the growth and development cycle are liable to reveal a range of larvae of slightly different ages. Thus, on any one day there may be three instars present. Alternatively, it was conceivable that the so-called brood 3 actually does not exist and what was observed is late brood 2 individuals and the onset of brood 4. This helped to explain the anomaly between Catling's statement that "Populations of yellow rice borer are always low before flooding (late June-early July)" and the fact that his light-trap catches showed no decline in numbers of adult moths at that time.
One of the difficulties that we faced in assessing the effectiveness of the insecticides used in the trials was that the numbers of live stages in the nontreated control appeared to decline with the passage of time, especially at Bastia (Table 1 - 1981). One reason may have been replacement of infested stems by new tillers, thus altering the stem population being sampled. This was reflected in the apparent drop in stem damage. Another, possibly more important, reason is that, unlike many other insect pests, which have much shorter generation times or less well-defined broods, or both, the yellow rice borer populations can increase only at the start of a fresh brood at four distinct points in each deepwater rice season.
Catling (1981) had given a good deal of information on moth numbers and egg parasitism. By combining his data with the report by Rothschild (1970) on the mortality of different stages from egg to pupa, we were able to derive a model of the yellow rice borer population. This is Figure 2.

The numbers of moths were taken from Catling's data from light trapping; the number of eggs per moth was taken as 300, which, like the level of egg parasitism, was derived from Catling's studies. The intermediate mortalities were from Rothschild (with some adjustment to fit the egg to adult figures). The gross mortalities for the broods are 95.6% for brood 2, 99.6% for brood 3, 98.3% for brood 4, and 99.6% for brood 5.
We stressed that importance should not be attached to the accuracy of those figures, nor should the numerical values for brood size be taken as indicative of actual field numbers. The importance was that the greatest decline in potential numbers comes between the laying of eggs and the detection of third-instar larvae. A high level of control is exerted by the environment and by natural enemies particularly in broods 3 to 5. Logistics, let alone environmental considerations, mitigate against use of insecticides after the onset of flooding.
From that, our thinking for the 1982 season was that the use of insecticides against brood 2 would be the only feasible time, and such use might depress the size of subsequent broods. Figure 2, if correct, showed that brood 2 is the only brood in which the degree of natural control could realistically be enhanced. The model showed also that those larvae that penetrate into the stem are protected from outside influences and, thus, mortality is low. It seemed possible, therefore, that one or two strategically timed applications of a suitable insecticide at the time of laying of the brood 2 eggs on deepwater rice seedlings might prove highly effective. Further applications at the time of laying of the brood 3 eggs, if not precluded by the onset of the floods, might be useful but the effect this would have on natural enemies would need to be established.
Before starting our studies, we hypothesized that if it was possible to reduce the preflood brood of yellow rice borer and, thus, depress the size of subsequent broods there could be a consequent reduction in stem infestation in what Catling (1981) described as the critical period, June to September.
In November, when reporting to the 1981 DWR conference, we felt that although there seemed little doubt that stem infestation is the best indicator of the numbers of stem borer larvae during June to September, it remained to be established whether or not such stem infestation is of prime importance in terms of actual crop loss. What if loss of the main and primary tillers is the major cause of yield reduction?
Setabutara & Vergara (1979) had found that, in one variety, 90% of the total yield came from the main and primary tillers combined; in a second variety, their contribution was 77%. Similarly, Datta & Banerji (1979) found that, regardless of water regimes, grain yield and yield-contributing factors of basal tillers in both the varieties they studied were always greater than those of aquatic tillers. About 70-80% of the total grain production came from basal tillers.
Catling (1981) reported that pot experiments showed a loss of 27-30% in panicle weight if basal tillers were lost and replaced by aquatic tillers. His crop loss studies in 1978 and 1979 with the variety Chota Bawalia showed that, although season-long insecticide use significantly reduced the stem infestation by yellow rice borer (as recorded at harvest) in both years and from identical levels the yield improvement was much less in 1979. The 1979 spraying of variety Lakshmidigha also gave a reduction in infestation but did not give a significant improvement in yield (in fact, only 3%).
My interpretation was that, not forgetting that 1979 was a poor year for crop establishment because of prolonged early drought, the major difference between the yield results in 1978 and 1979 might well have been that in the latter year yellow rice borer outbreaks did not occur until August and September. Therefore, in 1979 there could have been a lower than usual loss of basal tillers in the preflood period.
A re-examination of the field records of the 1978 study showed that no dissections were made in the early season but the deadheart level on 12 May was 14 times lower in the treated area than in the nontreated area (0.5/m² cf. 7.0/m²). There were few records, from 1977 to 1980, relating to yellow rice borer infestation before late June, but in 1978, at Agrakhola, deadhearts reached a peak of 11.0/m² on 19 May.
In 1981, at Bastia, our nonsprayed control plots gave the following results: (of 100 stems examined and dissected) 22 May, 18% deadhearts and 19% infested; 4 June, 4% deadhearts and 11% infested; 17 June, 2% deadhearts and 6% infested; I July, 3% deadhearts and 9% infested. Frame counts of tillers enabled us to estimate that there were 1.2 deadhearts/m² on 13 May and about 10.8 deadhearts/m² on 4 June.
From the limited data available, our feeling was that at the beginning of the season deepwater rice can be regarded the same as dryland rice and deadheart incidence will give a reasonable indication of infestation and damage by yellow rice borer. In the young plant, however, where there is little lumen development, the yellow rice borer larva is likely to cause much more damage to conducting tissues and to the apical meristem than in older plants. The elongated deepwater rice plant, with its massive stem lumens, probably provides a safe stable environment for the larva of the yellow rice borer, which feeds on the parenchyma tissue without damaging vascular tissue (Catling 1981).
Thus, we argued that at this stage the yellow rice borer may achieve the status of the near-perfect parasite-host relationship. If correct, this would help to explain why the number ofwhiteheads at harvest represents less than 20% of the stem infestations recorded in the late season (Catling & Islam 1979).
We ended our conference report by outlining our future strategy. By the end of the season, our experiments were likely to give a reasonable indication as to which insecticides are most likely to be suitable for further testing and we hoped the 1981 yield data from Bastia would add weight to the view that there is a distinct place for preflood use of insecticides.
| ©2000 - Brian Taylor CBiol FIBiol
FRES 11, Grazingfield, Wilford, Nottingham, NG11 7FN, U.K. Visiting Academic in the Department of Life Science, University of Nottingham |
href="\dwr\ysb3.htm"